Effet allélopathique d’une espèce native sur une plante exotique envahissante majeure en Europe

Florence Piola1 (enseignante-chercheuse) et la classe de 2nd GT de l’Institution Sévigné de Mme Joséphine Villain Sawadogo2 (la liste des élèves est mentionnée en fin d’article)
Article original/Original article: Christina, Mathias, Soraya Rouifed, Sara Puijalon, Félix Vallier, Guillaume Meiffren, Floriant Bellvert, et Florence Piola. « Allelopathic Effect of a Native Species on a Major Plant Invader in Europe ». The Science of Nature 102, no 3 (5 mars 2015): 12. https://doi.org/10.1007/s00114-015-1263-x.
Institution : 1Université Claude Bernard Lyon 1, LEHNA UMR 5023, CNRS, ENTPE, F-69622, Villeurbanne, France
2Institution Sévigné Compiègne, 20 rue de la Sous-Préfecture, 60200 Compiègne

Résumé :
Cet article a été écrit par 9 élèves de l’Institution Sévigné de Compiègne avec leur professeur de SVT à partir d’une étude menée par l’équipe du Docteure Piola et avec sa collaboration. Ils se sont intéressés à l’article portant sur l’effet allélopathique d’une espèce native Sambucus ebulus sur une plante exotique envahissante majeure en Europe Fallopia x bohemica faisant partie des Renouées asiatiques. Grâce à cette réécriture, les élèves ont pu apprendre comment l’effet allélopathique d’une plante native pouvait influencer une plante exotique envahissante.
Mots clés : allélopathie ; plante exotique envahissante ; plante native ; Fallopia x bohemica
I. Introduction
De nombreuses plantes exotiques envahissantes ne sont pas dominantes dans leur habitat naturel mais peuvent le devenir en étant introduites dans un nouvel environnement (Ridenour & Callaway, 2001). La compétition directe entre les plantes exotiques envahissantes et natives est un processus par lequel les espèces envahissantes peuvent remplacer les espèces natives. Ceci entraîne une modification des communautés représentant parfois une menace pour la biodiversité. Parmi les interactions directes, la compétition pour les ressources (utilisation de n’importe quelle ressource par un organisme qui en réduit la disponibilité pour un autre organisme) est souvent considérée comme le principal facteur limitant pour la colonisation par une espèce. Cependant, d’autres formes de compétition directe, comme l‘allélopathie, peuvent également avoir un impact significatif (Michalet et al., 2006).
L’allélopathie est définie comme un processus d’interaction chimique plante-plante ou plante-micro-organisme. Elle a des effets positifs ou négatifs, et est aussi activement impliquée dans le succès des invasions. Diverses études ont conduit à l’hypothèse que les plantes qui ont co-évolué avec une espèce allélopathique peuvent être moins sensibles aux composés avec lesquels elles ont évolué tandis que les espèces nouvellement exposées vont présenter une moindre résistance face à la nouvelle espèce introduite produisant ces composés allélopathiques jusqu’alors inconnus. Le succès des espèces exotiques envahissantes va donc être plus important face aux plantes natives.
Toutefois, cette théorie a pour corollaire que les espèces exotiques envahissantes qui n’ont pas co-évolué avec les espèces natives pourraient être sensibles aux composés toxiques produits par ces dernières. Leurs composés allélopathiques pourraient affecter la croissance des espèces exotiques et constitueraient une nouvelle méthode naturelle de contrôle. Mais, les effets allélopathiques des plantes natives sur les espèces exotiques ont rarement été étudiés. Fallopia x bohemica est une des espèces les plus envahissantes en Europe. Elle a la capacité de se disperser en utilisant deux types de propagules, les akènes (dispersion sexuée) et les rhizomes (dispersion végétative) ce qui la rend très difficile à contrôler. Il a récemment été démontré que Sambucus ebulus (Sureau hièble) pouvait limiter la colonisation et le développement des deux types de propagules de F. x bohemica. Cet effet pourrait s’expliquer par un effet allélopathique direct, par un accès limité à la lumière ou par une compétition pour les ressources. En effet, la plante S. ebulus est capable de coloniser le même lieu d’habitation (par exemple les bords de route ou les berges de rivière) que F. x bohemica. Nous pouvons supposer que les propagules sexuées et les propagules végétatives comme les fragments de rhizome sont impliquées dans la colonisation et la propagation pendant l’invasion.
Dans ce travail, il a été émis comme hypothèse que l’espèce native S. ebulus a un effet allélopathique sur l’espèce exotique envahissante F. x bohemica, limitant la croissance de celle-ci. Cette hypothèse a été testée avec des akènes et des fragments de rhizomes en utilisant un plan expérimental consistant à arroser les pots cibles de F. x bohemica avec les lessivats des pots donneurs de S. ebulus. L’effet des lessivats de S. ebulus sur les traits de croissance de Fallopia (hauteur, nombre de feuilles, masse sèche souterraine et masse sèche aérienne) a été mesuré.
II. Matériels et méthodes
1. Echantillonnage
| Date | Espèce | Mode de stockage et de nettoyage de culture | Lieu |
| 13/12/2010 | Rhizomes de S. ebulus collectés | 4°C Laver afin d’enlever la terre | Chambéon (France, Pays de la Loire) |
| Janvier/ Février 2011 | Rhizomes de F x bohemica collectés (2 échantillons de la même plante) | Cultiver à 20°C dans une serre dans des conditions identiques | Feyssine à Lyon => population 1 Veauchette (département de la Loire) -> population 2 |
| Janvier 2009 | Akène de F. x bohemica | Le long de la rivière Dorlay à Lorette (France) |
2. Plan expérimental
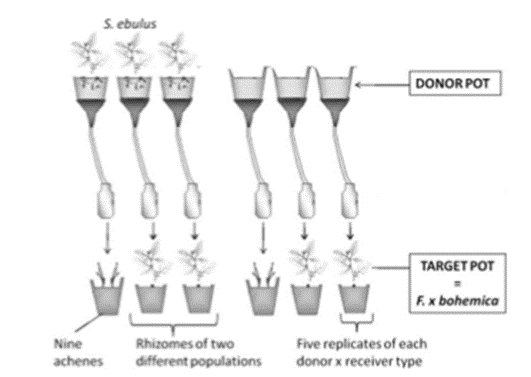

Le plan expérimental (Figure 1 et 2) a été adapté en utilisant des pots donneurs (pots contenants ou pas des plants de S. ebulus) et des pots cibles (avec ou sans akènes/rhizomes de Fallopia) pour séparer la compétition pour les ressources et l’allélopathie. Des pots de 1 L remplis de terreau Klasmann ont été utilisés. Le terreau permet de tester si une plante a un effet chimique direct sur une autre indépendamment des effets liés à la communauté microbienne, probablement peu représentée dans ce type de substrat.
Les pots donneurs ont été placés sur des entonnoirs en plastique de 15 cm de diamètre et ont été placés sur des étagères à 30 cm au-dessus des pots cibles. Un flacon de 120 mL a été placé sous chaque entonnoir pour recueillir les lessivats qui sont des solutions formées par le mouvement ou la percolation d’un liquide à travers un sol ou des déchets solides et la dissolution dans l’eau de certains constituants des milieux traversés. Trente pots ont été assignés au hasard aux contrôles (15 pots sans plantes) ou au traitement (15 pots contenant des rhizomes de S. ebulus). Les fragments de rhizomes ont été sélectionnés avec un nœud et une biomasse d’environ 5g±0,1 g.
Les rhizomes de S. ebulus avaient été plantés 2 mois avant le début de l’expérience afin d’obtenir un développement aérien de chaque rhizome. Des pots donneurs témoins (juste du sol) ont également été préparés et arrosés pendant ces 2 mois avant le début de l’expérience. Au début de l’expérience, les 30 pots donneurs ont été saturés d’eau et ensuite arrosés tous les 2 ou 3 jours, respectivement, pour obtenir 100 ml de lessivat.
Trente pots cibles contenaient des propagules de F. x bohemica : 10 avec des akènes (9 akènes par pot), 10 avec des fragments de rhizome de la population 1 et 10 avec des fragments de rhizome de la population 2 (1 fragment par pot). Les fragments de rhizome ont été sélectionnés avec un nœud et une biomasse de 1,5g± 0,1 g. Ces pots ont été régulièrement arrosés avec les lessivats des pots donneurs récoltés par gravité (pour chaque ensemble de 10 pots cibles, 5 ont été arrosés avec des lessivats de pots donneurs témoins et 5 avec les pots donneurs contenant des plantes de S. ebulus). Les pots cibles ont été déplacés aléatoirement chaque semaine au sein d’un même traitement (témoin ou S. ebulus) afin d’éviter un éventuel effet de localisation. L’expérience a duré 50 jours.
Explication sous forme de tiret pour simplifier la démarche :
- Utilisation pots donneurs et cibles, terreau Klasmann.
- Pots donneurs > entonnoirs 15 cm sur étagères 30 cm au-dessus pots cibles. + Flacon 120 mL sous entonnoir
- 15 pots contrôles : sans plantes ; 15 pots au traitement : rhizomes S ebulus (nœud et biomasse 5+0,1g)
- Plantation S. ebulus 2 mois avant expérience > pousses aériennes. Préparation et arrosage pots donneurs 2 mois avant expérience
- Saturation d’eau de 30 pots donneurs puis arrosage tous les 2 ou 3 jours respectivement > obtention 100 mL lessivats
- 30 pots cibles contenaient F. x bohemica, 10 avec akènes (9 par pots), 10 avec fragments rhizomes de population 1 et 10 avec fragments rhizomes de population 2 (1 fragment par pot) (nœud et biomasse de 1,5g+0,1g)
- Arrosage régulier des pots avec lessivats (pour chaque ensemble de 10 pots cibles, 5 arrosés avec lessivats pots donneurs, et 5 avec pots donneurs contenant S.ebulus)
- Déplacement aléatoire des pots cibles chaque semaine pour éviter un éventuel effet de localisation
- Durée de l’expérience : 50 jours
3. Mesures et analyses
Pour mesurer l’effet de S. ebulus sur Fallopia, le taux de germination des akènes et le temps régénération de rhizomes de F. x bohemica ont été évalués. Afin de tester l’impact des lessivats de S.ebulus sur la croissance de F. x bohemica , la hauteur, le nombre de feuilles et les masses sèches de celles-ci ont été mesurés durant 50 jours.
| Une analyse chimique de S.ebulus a été faite. Après broyage et extraction, le surnageant recueilli des rhizomes de S.ebulus par centrifugation, a été analysé parchromatographie liquide haute performance aussi appelé HPLC, dans le but de déterminer ses composants chimiques potentiellement allélopathiques. |
Toutes les analyses sont faites à l’aide du logiciel R 2.12.0 (R Development Core Team 2010), puis à l’aide d’ANOVA ou d’autres modèles mixtes d’analyse de variance.
III. Résultats
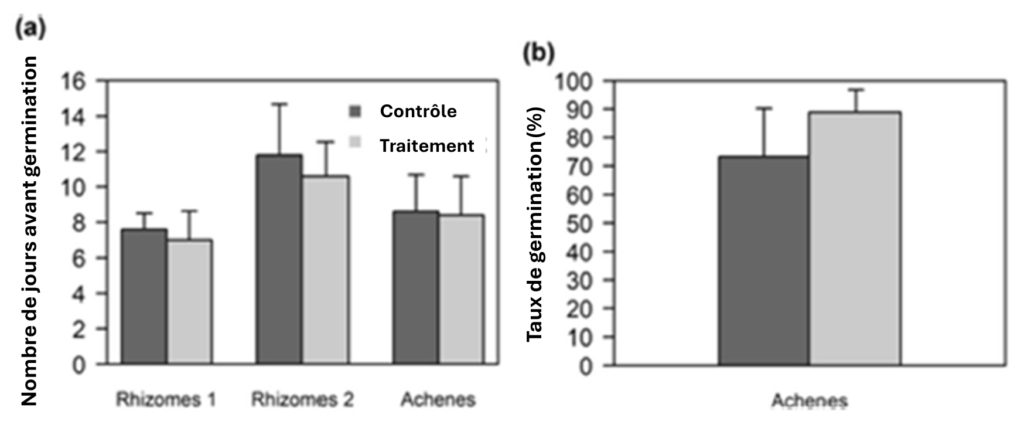
Le traitement au lessivat de S.ebulus n’a eu aucun effet sur la régénération de Fallopia comme le montre la figure 3 ci-dessous.Il n’a pas non plus eu d’effet sur le taux de germination des akènes.
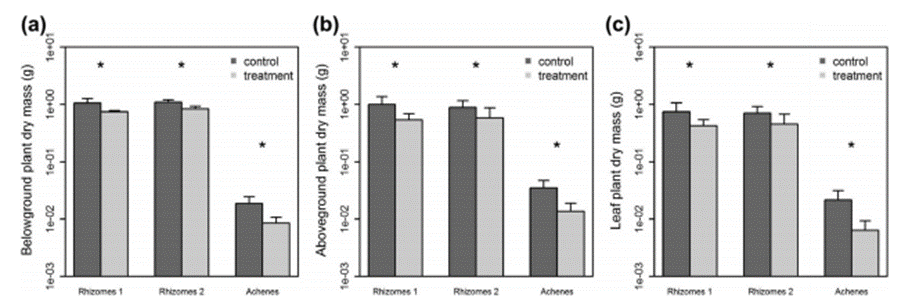
Dans le cas des plantes issues des akènes, le traitement a eu plusieurs effets sur les masses sèches (Figure 4). La masse sèche souterraine a diminué de 55%, la masse sèche aérienne a diminué de 61% et la masse sèche foliaire a, elle, diminué de 70% par rapport aux plants issus d’akène n’ayant pas reçu de lessivats de S. ebulus. La hauteur a aussi diminué, de 38% pour les akènes traités. Au bout de 50 jours, le traitement a diminué de 20% le nombre total des feuilles.
Dans le cas des plantes provenant des rhizomes, l’interaction entre le traitement et les populations de rhizomes n’était pas significative pour aucun des traits mesurés. Mais, le nombre de jours avant l’émergence, la hauteur des plantes et le nombre de feuilles étaient différents entre les plantes provenant de la population de rhizomes 1 et la population de rhizomes 2. Les plantes provenant de la population de rhizomes 2 étaient plus courtes, avaient moins de feuilles et prenaient plus de temps à émerger.
Le traitement a fait diminuer la masse sèche souterraine, la masse sèche aérienne, celle des feuilles ainsi que la hauteur des deux populations de rhizomes. La masse sèche souterraine a diminué de 25%, la masse sèche aérienne a diminué de 41% et la masse sèche des feuilles a diminué de 40%. La hauteur a aussi diminué de 31% pour les rhizomes 1 et de 26% pour les rhizomes 2 (Figure 4).
L’espèce introduite est sensible aux métabolites secondaires produits par l’espèce native (S.ebulus).
Quels sont les métabolites secondaires produits par S. ebulus ?
La chromatographie liquide haute performance à 280 nm de l’extraction des rhizomes de S.ebulus a montré sept pics principaux, identifiés comme étant des composés chimiques, des métabolites secondaires. Ce sont les flavonoïdes, la catéchine et l’épicatéchine, quatre autres ont montré des caractéristiques de la famille des acides di-hydroxycinnamiques et le dernier composé phénolique n’a pas pu être identifié.
IV. Discussion
Les expériences qui démontrent l’allélopathie sont souvent critiquées en raison de la difficulté à mettre en place un dispositif expérimental adéquat. Ici la méthode implique des pots cibles arrosés par des lessivats des pots donneurs pour étudier l’allélopathie. L’utilisation du dispositif a permis de valider l’hypothèse de l’implication de l’allélopathie dans les interactions entre les plantes natives et invasives. Dans une étude précédente, la croissance de Fallopia a été affectée par la compétition de S.ebulus. Cette compétition dans le même pot est probablement due à une interférence allélopathique.
F. x bohemica peut se propager végétativement et grâce à la reproduction sexuée. La dispersion potentielle des akènes de Fallopia le long des rivières a été mise en évidence (Rouifed et al., 2011). Dans cette étude, les plantules issues d’akènes semblaient beaucoup plus sensibles à S. ebulus que les plantules issues de rhizomes (Rouifed, 2011). Les résultats obtenus confirment l’hypothèse proposée. L’application de lessivats sur les akènes a entraîné une diminution plus importante de la hauteur, du nombre de feuilles ou de la masse sèche souterraine des plantes que sur les rhizomes. Cette sensibilité pourrait participer à la prédominance de la dispersion clonale.
Les lessivats de S. ebulus n’ont eu aucun effet sur l’émergence des plantes. Mais au cours du stade de développement il y a un impact sur la croissance, la hauteur ainsi que sur la biomasse sèche de Fallopia. L’allélopathie peut donc être impliquée dans la résistance à l’invasion. Dans certains fruits et certaines feuilles de Myrica gale, il y a un composé allélopathique, la myricagalone, qui est phytotoxique pour F.x bohemica (Popovici et al., 2011). On montre ici que F.x bohemica est aussi sensible aux composés chimiques provenant d’une plante native, dans des conditions plus proches des conditions naturelles.
Les interactions entre les populations de Fallopia et les populations de Sambucus pourraient faire évoluer les populations de Fallopia pour qu’elles tolèrent l’effet de la nouvelle allélochimie de la plante native. La chromatographie d’extraits de rhizomes de S. ebulus a révélé des composés connus pour être potentiellement allélopathiques (Macias et al., 2007) comme la catéchine est un composé chimique impliqué dans le succès invasif de C. maculosa (Thorpe et al., 2009).
Selon l’équipe de recherche, une façon d’identifier quels composés particuliers du S. ebulus sont impliqués dans la compétition contre Fallopia serait de compléter les pots cibles de F. x bohemica avec certains des métabolites secondaires clés identifiés dans le cocktail du S.ebulus. Ces métabolites peuvent également cibler les micro-organismes, et les effets allélopathiques observés peuvent résulter d’interactions indirectes impliquant des micro-organismes (Inderjit et al., 2011). Dans cette étude, même si des précautions ont été prises avec l’utilisation d’un terreau, les microorganismes présents dans les pots donneurs peuvent modifier le cocktail biochimique de S. ebulus. D’autres expériences sont alors nécessaires pour approfondir les mécanismes impliquant les micro-organismes, qu’ils soient les cibles du processus allélopathique ou des organismes interférents du sol.
En conclusion, il a été montré que F. x bohemica est sensible aux substances allélopathiques produites par l’espèce native S. ebulus. Ces résultats suggèrent un nouveau type de résistance biotique, peu étudié jusqu’à présent, suivant l’hypothèse inversée des nouvelles armes.
| Lexique : Allélopathie : Ensemble des interactions chimiques entre deux ou plusieurs plantes Espèce native :Espèce végétale ou animale qui vit dans son aire de répartition naturelle ou de dispersion potentielle. Propagules : Petit organe pluricellulaire (qui comporte plusieurs cellules) assurant la multiplication d’une espèce. ANOVA : Signifie l’analyse de la variance (Analysis of variance) Surnageant : Fraction la moins dense d’un liquide qui se trouve en surface. Rhizomes : Tiges souterraines de certaines plantes vivaces. Organes de multiplication végétative. Centrifugation : Procédé de séparation des composés d’un mélange en fonction de leur différence de densité en les soumettant à une force centrifuge. HPLC : La chromatographie en phase liquide à haute performance (HPLC) est une technique de chimie analytique utilisée pour séparer les composés d’un mélange chimique. |
Références :
Inderjit, Wardle, D. A., Karban, R., & Callaway, R. M. (2011). The ecosystem and evolutionary contexts of allelopathy. Trends in Ecology & Evolution, 26(12), 655‑662. https://doi.org/10.1016/j.tree.2011.08.003
Macias, F. A., Molinillo, J. M., Varela, R. M., & Galindo, J. C. (2007). Allelopathy—A natural alternative for weed control. Pest Management Science: Formerly Pesticide Science, 63(4), 327‑348.
Michalet, R., Brooker, R. W., Cavieres, L. A., Kikvidze, Z., Lortie, C. J., Pugnaire, F. I., Valiente-Banuet, A., & Callaway, R. M. (2006). Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecology Letters, 9(7), 767‑773.
Popovici, J., Bertrand, C., Jacquemoud, D., Bellvert, F., Fernandez, M. P., Comte, G., & Piola, F. (2011). An allelochemical from Myrica gale with strong phytotoxic activity against highly invasive Fallopia x bohemica taxa. Molecules, 16(3), 2323‑2333.
Ridenour, W. M., & Callaway, R. M. (2001). The relative importance of allelopathy in interference : The effects of an invasive weed on a native bunchgrass. Oecologia, 126, 444‑450.
Rouifed, S. (2011). Bases scientifiques pour un contrôle des renouées asiatiques : Performances du complexe hybride Fallopia en réponse aux contraintes environnementales.
Rouifed, S., Puijalon, S., Viricel, M.-R., & Piola, F. (2011). Achene buoyancy and germinability of the terrestrial invasive Fallopia× bohemica in aquatic environment : A new vector of dispersion? Ecoscience, 18(1), 79‑84.
Thorpe, A. S., Thelen, G. C., Diaconu, A., & Callaway, R. M. (2009). Root exudate is allelopathic in invaded community but not in native community : Field evidence for the novel weapons hypothesis. Journal of Ecology, 641‑645.
Ont participé au travail d’écriture de cet article, en collaboration avec Florence Piola, enseignante chercheuse en écologie (par ordre alphabétique) : BAVART Noah, BITULI Laura, BOULET Ilona, FAUX Flavie, GRAMONT Paul, POULAIN-RABIER Clémence, QUIRIN Anne-Flore, LE DEVEHAT Mathéo, MINOS Ludmilla.
Comment citer cet article : Florence Piola et la 2nde GT de l’Institution Sévigné (Compiègne, FR), Effet allélopathique d’une espèce native sur une plante exotique envahissante majeure en Europe, Journal DECODER, 2024-02-01