Les dinosaures couvaient-ils leurs œufs ?

Romain Amoit1 (enseignant-chercheur) et la classe de 2nd 9 du lycée Pierre Brossolette de Mme Christine Louis2 et Mr Odile Descos2 (la liste des élèves est mentionnée en fin d’article)
Article original/Original article: Bi, Shundong, Romain Amiot, Claire Peyre de Fabrègues, Michael Pittman, Matthew C. Lamanna, Yilun Yu, Congyu Yu, et al. « An oviraptorid preserved atop an embryo-bearing egg clutch sheds light on the reproductive biology of non-avialan theropod dinosaurs ». Science Bulletin 66, no 9 (15 mai 2021): 94754. https://doi.org/10.1016/j.scib.2020.12.018.
Institution : 1 CNRS UMR 5276, Laboratoire de Géologie de Lyon, Terre, Planètes, Environnement, 2 rue Raphaël Dubois, 69622 Villeurbanne Cedex.
2Lycée Pierre Brossolette, 1 Esplanade Agnès Varda, 69100 Villeurbanne

Résumé :
L’origine de la biologie de reproduction des oiseaux actuels pose beaucoup de questions. Ce questionnement est dû à la découverte de similitudes avec les dinosaures non aviens, notamment au niveau de la couvaison. La découverte d’un spécimen fossile dans le sud de la Chine apporte des éléments nouveaux à la compréhension des stratégies de reproduction des dinosaures non aviens.
Le spécimen étudié est un squelette de théropode adulte fossilisé en position accroupie au sommet d’une ponte de 24 œufs.
Des analyses phylogénétiques, histologiques et isotopiques ont été réalisées. Les résultats confirment qu’il s’agit d’un individu fossilisé en posture de couvaison. Les études ontogénétiques indiquent que c’est un adulte. Quant au sexe, il n’est pas déterminé.
Les œufs trouvés dans le nid contiennent des embryons à différents stades de développement, suggérant une stratégie d’éclosion asynchrone. La température d’incubation chez ces théropodes non aviens correspond à celle des dinosaures aviens.
Le résultat de toutes ces analyses atteste bien l’hypothèse que la reproduction des oiseaux n’est pas issue d’un processus évolutif linéaire, mais que diverses caractéristiques liées à la reproduction avienne sont apparues indépendamment au sein de leurs ancêtres dinosauriens. D’autres études viennent confirmer que l’évolution de la biologie de reproduction des oiseaux modernes s’est faite progressivement et certaines caractéristiques sont issues des théropodes non aviens.
Mots clés : oviraptorosaures ; couvaison ; éclosion asynchrone ; delta 18O ; phylogénie
I. Introduction
La biologie de reproduction des oiseaux modernes diffère de celle des autres vertébrés actuels, et son évolution a suscité beaucoup d’études. Une découverte clé liée à l’évolution de la biologie de reproduction des oiseaux modernes est le comportement de couvaison identifié chez les dinosaures non aviens par plusieurs spécimens. Ces derniers sont des squelettes d’oviraptorosaures préservés accroupis sur des amas d’œufs découverts en Chine et en Mongolie. L’interprétation d’une couvaison pour ces spécimens adultes associés à la ponte représente l’hypothèse privilégiée, mais d’autres interprétations valables existent également, y compris celles selon lesquelles le parent a péri en train de pondre ou de protéger ses œufs. Il n’existe aucune preuve rejetant de manière indiscutable la mort de l’adulte pendant la ponte. Plusieurs indices montrent que la mort de la femelle pendant la ponte est plus vraisemblable que lors de la couvaison. En effet, il existe des différences significatives de taille entre les différentes pontes découvertes. Ces différences pourraient correspondre à différentes étapes de la construction du nid. L’autre argument majeur privilégiant la mort de la femelle pendant la ponte est le contact limité entre les œufs et l’adulte. Nous décrivons ici un nouveau spécimen qui renforce non seulement l’hypothèse de la couvaison pour les oviraptorosaures, et qui fournit également des informations sur la biologie de reproduction des théropodes non aviens et d’autres groupes de théropodes.
II. Matériels et méthodes
1. Matériels
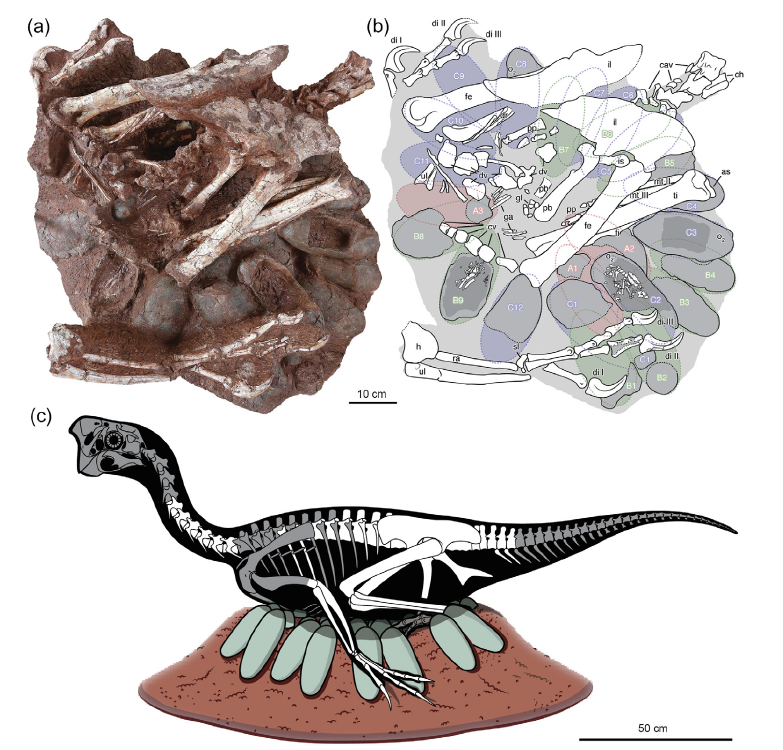
Le spécimen LDNHMF2008 (conservé au Lande Museum of Natural History, Tangshan, province du Hebei, Chine) est un squelette partiellement articulé d’un théropode de la famille des Oviraptoridae de taille moyenne auquel il manque le crâne, une partie des vertèbres cervicales, d’une grande partie des vertèbres dorsales et caudales, des ceintures pectorales et une majeure partie des humérus, préservé au sommet d’une ponte d’au moins 24 œufs non perturbés, dont certains sont cassés, laissant apparaitre des os embryonnaires (Fig. 1). Le spécimen a été découvert dans le Crétacé supérieur (Maastrichtien ; entre -72 et -66 MA) de la formation de Nanxiong près de la gare de Ganzhou, dans la province du Jiangxi en Chine.
2. Analyses histologiques
Des échantillons de coquilles d’œufs ont été prélevés sur les œufs C2, C5 et C8 (Fig. 1), tous préparés sous forme de lames minces pétrographiques et étudiés au microscope à lumière polarisée. Un gastrialium et l’humérus de l’adulte ont également été échantillonnés pour la préparation de lames minces ostéohistologiques, qui ont été étudiées au microscope à lumière polarisée pour examiner l’histologie osseuse.
L’analyse histologique est une méthode utilisée en biologie, en médecine et en paléontologie pour étudier la structure microscopique des tissus biologiques. L’histologie est la discipline qui consiste à prélever des échantillons de tissus (généralement à partir de biopsies ou de prélèvements post-mortem), à les traiter, à les couper en fines sections, à les colorer avec des colorants spécifiques, et à les examiner au microscope pour étudier leur composition cellulaire, leur organisation et leur relation avec d’autres tissus osseux
3. Analyses phylogénétiques
La phylogénie est l’étude des liens de parenté entre différentes espèces. Elle est souvent représentée sous formes d’arbres pour montrer les relations entre des espèces partageant un ancêtre commun (Fig. 2).
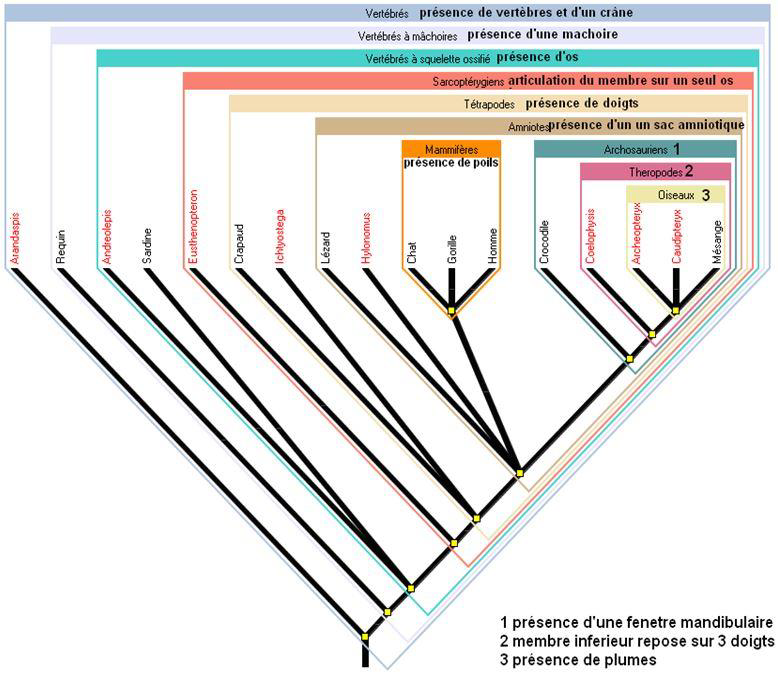
Chaque ancêtre commun partage des caractéristiques anatomiques propres avec les espèces issues de cet ancêtre commun.
Un caractère qui ne subit pas de modification au cours de l’évolution est appelé caractère plésiomorphe ou caractère ancestral. C’est le cas de la ponte synchrone (ponte au sein de laquelle tous les œufs se développent et éclosent en même temps).
Un caractère résultant d’une modification au cours de l’évolution est appelé caractère dérivé. C’est le cas de la ponte asynchrone (Ponte au sein de laquelle les œufs n ‘éclosent pas tous en même temps mais sur une période). Il s’agit d’une modification du type de ponte au cours de l’évolution.
La position systématique du nouveau spécimen a été déterminée grâce à deux analyses distinctes en utilisant un logiciel d’analyse phylogénétique. Tout d’abord une analyse phylogénétique basée sur un ensemble de données de théropodes coelurosauriens récemment publié avec l’ajout de LDNHMF2008 a été réalisée. Une deuxième analyse phylogénétique a été réalisée en utilisant un ensemble de données récemment publié sur la phylogénie des oviraptorosaures avec l’ajout de LDNHMF2008.
4. Analyses isotopiques
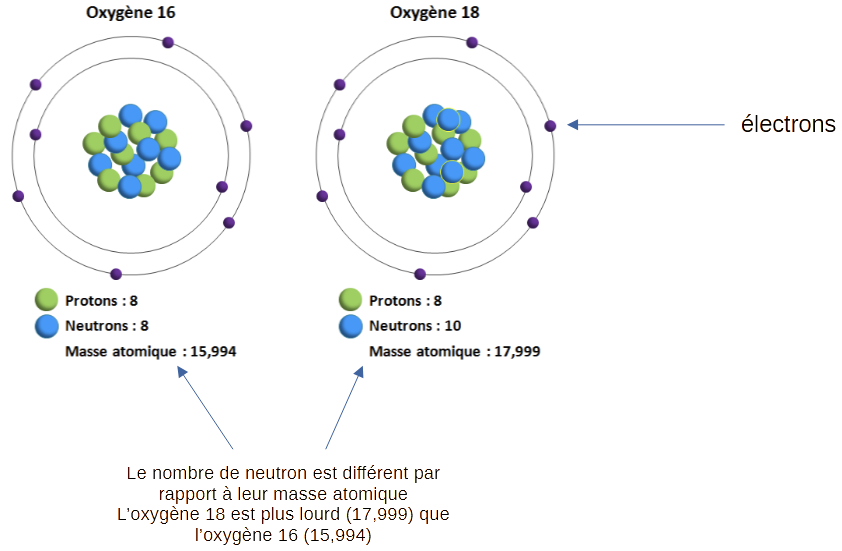
Les isotopes sont des atomes qui possèdent un même nombre d’électrons et donc de protons pour rester neutres, mais un nombre différent de neutrons (Fig.3).
III. Résultats
Le spécimen LDNHMF2008 présente la posture de couvaison in situ typique observée sur d’autres oviraptorosaures préservés au sommet de pontes d’œufs (Citipati osmolskae : IGM 100/979, IGM 100/1004 ; Nemegtomaia barsboldi : MPC-D 107/15 ; Oviraptoridae indet. : IVPP V9608) : les membres antérieurs sont orientés ventralement et postérieurement (c’est-à-dire vers le bas et vers l’arrière), recouvrant les bords de la ponte ; les membres postérieurs sont repliés de telle manière que le corps de l’animal est situé au-dessus des œufs et centré sur la ponte. La région pelvienne repose en partie sur la partie postérieure de la ponte et en partie au centre. Le corps est préservé à une proximité extrêmement étroite des œufs (Fig. 1).
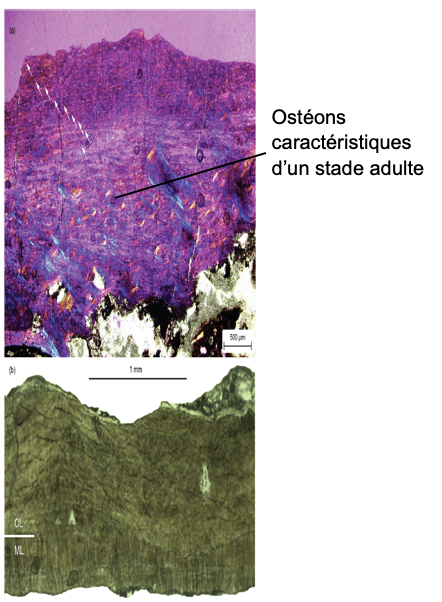
Le spécimen est vraisemblablement un adulte, comme l’indique la structure anatomique des vertèbres observées. Une analyse du tissu osseux a été réalisée pour en déterminer davantage sur son stade de développement. Pour ce faire, des lames minces d’os sont observées (Fig. 4a). Les observations réalisées ont permis de conclure que l’individu étudié était un adulte.
Les lames minces montrent clairement l’absence d’os médullaire dans l’humérus* et le gastrialium*. L’os médullaire est produit exclusivement par la femelle pendant une période et sert de stockage de calcium qui est ensuite utilisé pour la minéralisation des coquilles d’œufs. On peut supposer que l’individu est un mâle car il n’a pas d’os médullaire mais on ne peut pas totalement exclure que ce soit une femelle.
Le squelette adulte présente des caractéristiques anatomiques générales typiques des oviraptorosaures, de plus, le spécimen LDNHMF2008 a été identifié comme faisant partie de la famille des Oviraptoridae. Cette détermination a été obtenue grâce aux analyses phylogénétiques basées sur l’étude des caractères présents sur le bassin, le fémur et les os du pied de ce dinosaure. Une autre analyse osseuse basée sur les vertèbres caudales et le pubis a donné des résultats différents, indiquant une position au sein de Caenagnathoidea. (Fig. 5).

Figure. 5 : Position phylogénétique de LDNHMF2008, Arbre simplifié d’après la figure originale de l’article. Les images sont extraites : https://www.anyrgb.com/; Zhao Chuang ;Wikipedia ; https://fossil.fandom.com/
La ponte se compose d’au moins 24 œufs conservés immédiatement sous et en proximité extrêmement étroite du squelette de l’adulte, avec peu ou pas de matrice sédimentaire entre les deux. Les diamètres externe et interne de l’ensemble de la ponte sont respectivement de 78 et 26 cm. La ponte est principalement intacte. Elle ne présente qu’une légère déformation ou compression des œufs. Par ailleurs, il y a conservation de l’appariement des œufs qui sont disposés en trois cercles superposés. En effet, douze œufs sont exposés dans le cercle supérieur (cercle C), neuf dans le cercle du milieu (B), et trois dans le cercle inférieur (A). Le nombre total d’œufs est supérieur à celui rapporté dans d’autres pontes associées à des adultes oviraptoridae (de 6 à 22 œufs par ponte).
Les œufs mesurent environ 215 mm de long et 85 mm de large (Tableau S1 en ligne). Chacune des coquilles présente une ornementation composée de fines crêtes mesurant environ 2 à 3 mm de largeur. Les crêtes sont alignées avec l’axe vertical des œufs et sont disposées de manière compacte vers son équateur mais de façons plus disparates à ses deux extrémités. La coquille d’œuf est fine, d’environ 1,08 mm d’épaisseur sans prendre en compte la crête. La coquille d’œuf est formée d’une couche dite mamillaire interne séparée par une limite ondulée surmontée d’une couche continue (Fig 4b). Grâce à la taille des œufs, à l’épaisseur de leur coquille et celle de la limite ondulée qui est distincte entre les deux autres couches de la coquille d’œuf, les œufs sont attribués à l’espèce Macroolithus yaotunensis.

Dans sept œufs il est possible d’observer un squelette d’embryon. Le squelette de l’œuf C2 est partiellement articulé et montre une posture recroquevillée (Fig. 6b), comme dans d’autres embryons d’oviraptorosaures. Son stade d’ossification est similaire à celui observé dans les toutes dernières étapes du développement des embryons d’oiseaux du groupe couronne* et d’autres oviraptorosaures. Les différences de taille et de stades de développement entre ces embryons peuvent être une conséquence de l’éclosion asynchrone, comme cela a été suggéré précédemment pour les oviraptorosaures.
L’œuf B9 (Fig. 6a) présente moins d’éléments apparents que les œufs de la couche C. La plupart des os dans l’œuf B9 ne peuvent pas être identifiés. L’embryon dans l’œuf B9 est considéré comme étant plus précoce sur le plan du développement que ceux des œufs C2 et C5. En général, une variation de développement est observée non seulement entre les œufs de différents cercles, mais aussi à l’intérieur du même cercle.
Pour estimer la température à laquelle les embryons ont minéralisé leurs squelettes, les compositions isotopiques de l’oxygène du phosphate de l’hydroxyapatite des os embryonnaires et du carbonate de la coquille des œufs ont été mesurées et analysées (œufs analysés : C2, C3, C8). Cette mesure consiste à déterminer la proportion de 18O et de 16O dans la coquille et dans les os.
Avant d’exploiter les mesures, rappelons ce qu’est un os et où est situé l’atome d’oxygène étudié. Un os est une structure rigide constitué de composants organiques et inorganiques.
Les composants organiques sont des cellules spécialisées dans la fabrication de l’os.
Les composants inorganiques constituent 65 % de la masse de l’os. Son composant principal est un sel minéral de phosphate de calcium appelé hydroxyapatite (Ca10(PO4)6OH2). On étudie la proportion de 18O et de 16O de l’oxygène associé au phosphate (P) de l’hydroxyapatite.
Le calcul de la température d’incubation est basé sur les considérations suivantes :
La composition isotopique de l’oxygène de la coquille d’œuf reflète la composition isotopique de l’oxygène des fluides corporels de la femelle à partir desquels la coquille d’œuf précipite. Étant donné que l’eau des fluides de l’œuf provient de l’eau corporelle de la femelle et est transmise sans fractionnement isotopique, les valeurs isotopiques de la coquille reflètent également indirectement les compositions isotopiques de l’oxygène des fluides de l’œuf.
Les os des embryons se forment à partir des fluides de l’œuf, avec un fractionnement isotopique influencé par la température de l’œuf. Pour cela, nous avons utilisé une modélisation qui a permis de prédire l’évolution de la composition isotopique de l’oxygène du squelette des embryons pendant que le dinosaure couvait les œufs, en tenant compte de leurs températures respectives. Les valeurs mesurées de δ18O de la coquille d’œuf et des os des embryons ont été utilisées comme conditions limites* pour cette modélisation.
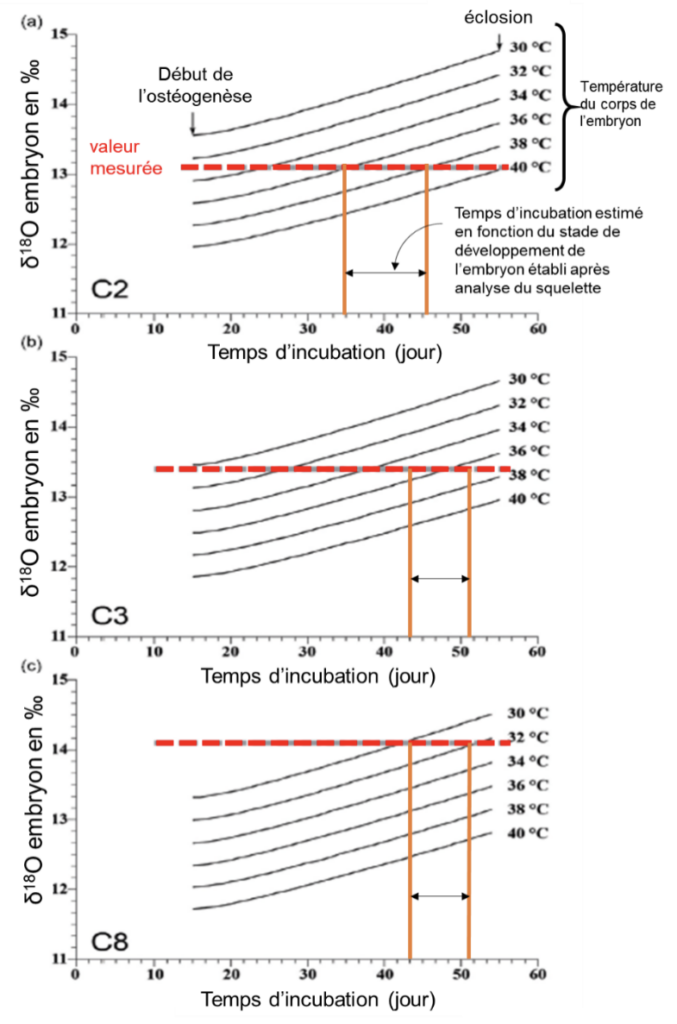
Ainsi, l’analyse isotopique montre que des différences de température existaient entre les œufs. Leur température pouvait varier de 30°C à 40°C. Les dinosaures non aviens avaient des températures corporelles élevées situées entre 33–38 °C. Deux des trois œufs analysés et couvés se situent bien dans la gamme de température attendue d’une couvaison (36–38 °C), tandis que le troisième embryon a été incubé à une température plus basse de 30–32 °C mais toujours dans la gamme des températures de couvaison.
Cette température inférieure calculée pourrait être le résultat d’une partie de la ponte qui aurait été moins couverte par le corps de l’adulte et donc avoir été soumis à une température plus basse.
On suppose que le stade de l’embryon dans l’œuf C8 serait légèrement plus précoce par rapport à ceux dans C2 ou C3. La température d’incubation plus basse à laquelle cet œuf a été soumis, a très certainement affecté la vitesse de développement embryonnaire et donc sa durée.
IV. Discussion
Le fossile LDNHMF2008 représente le premier spécimen de dinosaure non avien à conserver en association un adulte au sommet d’un nid d’œufs contenant des embryons. Étant donné que les embryons sont morts à des stades de développement avancés, l’adulte a clairement pris soin du nid pendant une période prolongée et n’est pas mort en pondant ses œufs, contrairement à ce qui a été suggéré précédemment pour d’autres découvertes.
La température d’incubation estimée des œufs est cohérente avec une température corporelle d‘oiseaux couveurs. En revanche, la température d’incubation des reptiles est toujours plus basse. Dans l’ensemble, la combinaison entre contact étroit entre le squelette adulte et les œufs, les stades de développement avancés des embryons et la température d’incubation estimée élevée exclut à la fois l’hypothèse de la mort de l’adulte pendant la ponte et celle de garde du nid sans couvaison comme cela est observé chez les reptiles actuels. Ces éléments soutiennent fortement l’hypothèse de couvaison pour les spécimens associant des oviraptorosaures (dinosaures proches des oiseaux) adultes à leurs pontes.
Des études précédentes démontrent que l’évolution de la reproduction des oiseaux modernes s’est produite en plusieurs étapes, et certaines caractéristiques de reproduction des oiseaux trouvent leurs origines au sein des théropodes non aviens, alors que d’autres caractéristiques ont évolué parmi les oiseaux primitifs. Les maniraptoriens du grade des oviraptorosaures représentent un stade intermédiaire dans l’évolution de la biologie de reproduction des oiseaux modernes, car la communauté scientifique considère qu’ils possèdent des caractéristiques telles que pondre en plusieurs fois, peut-être une pratique de couvaison paternelle pour aider à l’incubation dans un nid partiellement ouvert, et l’éclosion en même temps des œufs via une incubation retardée. Les découvertes rapportées ici soutiennent la présence de certaines (par exemple, la ponte en plusieurs fois et le comportement de couvaison paternelle) mais pas toutes (par exemple, l’éclosion en même temps des oeufs) de ces caractéristiques chez les oviraptorosaures.
Chez la plupart des oiseaux actuels nidifuges*, les embryons d’une même couvée se développent et éclosent en même temps, ou presque. L’éclosion synchrone est une caractéristique ancestrale (plésiomorphique*) pour les oiseaux du groupe couronne* et les autres vertébrés. En même temps les oiseaux nidicoles* sont caractérisés par une éclosion asynchrone, une caractéristique dérivée du groupe couronne. LDNHMF2008 présente un développement en une seule couvée. La différence de maturité développementale entre les œufs renforce l’idée que l’éclosion asynchrone aurait pu exister chez les théropodes non aviens. L’éclosion asynchrone est due à la différence de température entre les œufs/embryons au sein d’une seul couvée. Cependant, on ne peut pas exclure complètement que des échecs de développement de certains œufs pourraient avoir entrainé l’apparente variation des différents stades de développement embryonnaire observée dans la couvée.
En raison de la complexité des données sur LDNHMF2008 il a été impossible d’établir un schéma explicatif du développement des embryons donc nous ne sommes pas capables d’expliquer la cause de l’éclosion asynchrone*. Néanmoins il est possible de proposer une théorie avec les données disponibles sur l’éclosion asynchrone chez les oviraptosaures semblable. Comme chez les autruches, les oviraptorosaures commence l’incubation* des œufs une fois tous les œufs pondus.
Ainsi les œufs situés au niveau inférieur (le plus bas) et donc pondus en premier auraient eu un temps d’incubation proportionnellement équivalent aux œufs supérieurs, pondus plus tardivement.
Toutefois, les œufs supérieurs auraient éclos avant les œufs inférieurs vu qu’ils étaient plus proche du couveur. En effet, la température de couvaison étant plus élevée cela aurait accéléré le développement des embryons à l’intérieur. Du fait de cette distance variable entre les différents cercles d’œufs pondus, on peut supposer que les œufs auraient pu éclore à des moments différents.
La découverte de l’éclosion asynchrone chez les oviraptorosaures est inattendue compte tenu de son apparition tardive même parmi les oiseaux du groupe couronne. Apparemment, cette caractéristique a évolué indépendamment chez certains oiseaux du groupe couronne et au moins chez les oviraptorosauriens parmi les dinosaures non aviens, renforçant les hypothèses précédentes selon lesquelles l’évolution de la biologie de reproduction des oiseaux modernes n’était pas un processus linéaire. Au contraire, l’évolution indépendante de l’éclosion asynchrone chez un théropode non avien proche des oiseaux fournit un autre exemple de l’apparition d’un caractère non hérité d’un ancêtre commun à travers cette importante transition évolutive. Cette découverte met en lumière la complexité de l’évolution et la diversité des trajectoires évolutives possibles.
Les théropodes non aviens n’étaient pas, d’un point de vue comportemental, strictement intermédiaires entre les oiseaux et les autres archosaures, soulignant davantage l’unicité des différents sous-groupes de dinosaures non aviens. Enfin, le scénario que nous proposons pour l’éclosion des oviraptorosaures n’est pas entièrement cohérent avec les données des oiseaux actuels. Ils affichent normalement une éclosion synchrone si l’incubation commence après que tous les œufs d’une couvée ont été pondus. Cela met en évidence d’avantage les comportements complexes des dinosaures non aviens en tant que groupe d’animaux ayant une histoire évolutive propre et unique.
| LEXIQUE : Asynchrone : qui n’est pas en même temps Aviens : qui a des traits semblables aux oiseaux Caenagnathidae : famille de dinosaures ressemblant à des oiseaux. Ils sont éteints et vivaient en Amérique du Nord et en Asie au Crétacé. Conditions limites: Données le plus souvent mesurées qui permettent de limiter la modélisation à des résultats réalistes. Crétacé : Qui correspond à une période de la fin de l’Ere Mésozoïque entre -145 et -66 millions d’années. Eclosion : ouverture à maturité d’une enveloppe protectrice contenant un être vivant Gastrialium : plaques osseuses dermiques des reptiles. Les gastralia (singulier gastralium) sont des plaques osseuses dermiques des reptiles comme les crocodiliens, les Sphenodons, les squamates, les dinosaures et d’une certaine manière, les tortues1. On trouve ces plastrons ventraux chez l’archéoptéryx, mais pas chez les oiseaux actuels.Ces plaques de protection soutiennent les viscères et sont un pendant à la cage thoracique. Elles jouent donc un rôle dans la respiration. Elles peuvent également protéger les viscères des animaux qui en sont pourvus s’ils marchent en rampant sur le sol. Groupe couronne : petit clade formé par le dernier ancêtre commun. Humérus : os situé dans le bras, entre l’épaule et le coude Incubation : action de couver des œufs Nidicole : jeune qui reste au près du parent pour être nourri pendant les premières semaines Nidifuges : les jeunes oiseaux qui quittent le nid après l’éclosion Oiseaux nidicoles : Une espèce animale est dite nidicole ou altriciale lorsque ses petits naissent incapables de se nourrir et de se déplacer seuls. Généralement ils naissent nus et aveugles. C’est le cas notamment des pigeons et de nombreux passereaux. Ontogénique : Se dit des différents stades du développement d’un être vivant depuis sa conception jusqu’à l’état adulte Oviraptorosaures : dinosaures à plumes, semblables aux oiseaux, qui vivaient en Amérique du Nord et en Asie. Phylogénie : l’étude des liens de parenté entre les êtres vivants et ceux qui ont disparu. Plésiomorphique : état ancestral du caractère biologique étudié. Pleurocèle : Chez les dinosaures, dépressions ou cavités aériennes trouvées dans certains os, notamment les vertèbres, de certains dinosaures et qui auraient pu jouer un rôle analogue aux sacs aériens des oiseaux actuels Synchrone : qui se réalise en même temps Théropodes : Dinosaures appartenant au groupe de saurischiens, bipèdes et pour certains carnivores, aux membres antérieurs courts mais pourvus de griffes, aux membres postérieurs adaptés à la course. |
Référence :
Bi, S., Amiot, R., Peyre de Fabrègues, C., Pittman, M., Lamanna, M. C., Yu, Y., Yu, C., Yang, T., Zhang, S., Zhao, Q., & Xu, X. (2021). An oviraptorid preserved atop an embryo-bearing egg clutch sheds light on the reproductive biology of non-avialan theropod dinosaurs. Science Bulletin, 66(9), 947‑954. https://doi.org/10.1016/j.scib.2020.12.018
Ont participé au travail d’écriture de cet article, en collaboration avec Romain Amiot, chercheur en paléontologie (par ordre alphabétique) : AATOUT Najib, ABDELADIM Mahera, ABDESSELEM Iheb-Israfil, AISSA Aya, AOUANI Ayoub, AUDEL Yla, AYOUAZ Hafsa, AZOUAOU Aimad, BOUHADJELA Camelia, BOULEMAIZET Mehedine, BOULENDJAS Méliha, BOUTCHICHA Kays, BRESSET–MORRY Ambre, BUTTAY Marion, DEBBAH Morjane, DELAVAUD–BURNAZ Marius, FAYE Penda, FRAHTI Waris, GERIN GUERICHE Faustine, LE JEUNE Paul, LETIFI Djihane, MECHERI Sarah, MHADHBI Wael, MOKDAD Noham, MOREIRA LEAL Ana, MOREL Arthur, MUKWABANGO Cynthia, OUKADIR Sara, OZCAN Elif, POZZI Ornella, SEFIANE Amani, SMAIL Samy, SWOROWSKI Samuel, TEZO AKUE Anne Laurie
Comment citer cet article : Romain Amiot et la 2nde 9 du lycée Pierre Brossolette (Villeurbanne, FR), Les dinosaures couvaient-ils leurs œufs ? Journal DECODER, 2024-06-27